Ismail G Said1,
Nayera AM Abdelwahed1,
Hassan M Awad1,2 ![]() ,
Madgy A Shallan1,
Kamal YI El-Shahed1,
Emam A Abdel-Rahim3
,
Madgy A Shallan1,
Kamal YI El-Shahed1,
Emam A Abdel-Rahim3
For correspondence:- Hassan Awad Email: awadmhassan@yahoo.com
Received: 6 September 2016 Accepted: 19 December 2016 Published: 31 January 2017
Citation: Said IG, Abdelwahed NA, Awad HM, Shallan MA, El-Shahed KY, Abdel-Rahim EA. Enhancement of clavulanic acid production by Streptomyces sp MU-NRC77 via mutation and medium optimization. Trop J Pharm Res 2017; 16(1):31-42 doi: 10.4314/tjpr.v16i1.5
© 2017 The authors.
This is an Open Access article that uses a funding model which does not charge readers or their institutions for access and distributed under the terms of the Creative Commons Attribution License (http://creativecommons.org/licenses/by/4.0) and the Budapest Open Access Initiative (http://www.budapestopenaccessinitiative.org/read), which permit unrestricted use, distribution, and reproduction in any medium, provided the original work is properly credited..
Purpose: To enhance clavulanic acid production using UV-mutagenesis on Streptomyces sp. NRC77.
Methods: UV-mutagenesis was used to study the effect of Streptomyces sp. NRC77 on CA production. Phenotypic and genotypic identification methods of the promising mutant strain were characterized. Optimization of the fermentation medium and culture conditions were investigated
Results: Out of the screened mutants, 120A3 mutant isolate was selected as promising. The phenotypic properties of 120A3 mutant showed culture characteristics similar to those of Streptomyces species. Phylogenetic analysis of 16S rRNA gene sequence indicate that this strain has similarity (99 %) to Streptomyces sp.T2-7; therefore it was suggested as Streptomyces sp. MU-NRC77 and has Gen Bank accession no. KT953342.
Conclusion: Improvement of CA yield by 48 % was obtained from fermentation medium and culture condition optimization. Further optimization by addition of H2O2 and activated charcoal to the production medium increased CA yield to 646.12 and 682.94 mg/L respectively, i.e., 83 % more than that obtained prior to addition.
Introduction
β-lactam ring-containing molecules such as those of penicillins and cephalosporins were hydrolyzed by the effect of the catalytic ability of enzymes produced by many pathogenic bacteria, resulting in the blocking of their antibacterial property, in turn causing resistance to antibiotics and creating a problem in the treatment of infectious diseases [1]. Clavulanic acid (CA) is a potent ß-Lactamase inhibitor that has been successfully employed in clinical practice. It is produced by the actinomycete Streptomyces clavuligerus, and is used in combination with beta-lactam antibiotics that are sensitive to attack by beta-lactamase enzymes. It acts by specifically inhibiting the active centers of these enzymes, hence avoiding loss of the beta-lactam antibiotic and restoring its antimicrobial action. The combined action as ß-lactamase inhibitor and antibacterial agent makes CA very important both clinically and economically [2].
Media composition, growth conditions optimization and strain improvement increased the production of CA (up to approx. 1.4 g/l) [3]. Strain improvement was achieved by mutagenesis, the most generally utilized tool for better yield of secondary metabolites in the fermentation process [4]. A relatively safe mutagen like ultraviolet (UV) irradiation represents a very convenient system to enhance strains through random mutation. But only a small number of researchers have utilized UV irradiation to overproduce clavulanic acid [3]. The taxonomic properties of S. olivaceus, such as the color of the aerial mycelium, liquefaction of gelatin, growth of cellulose, production of ammonia and nitrate reduction were changed after irradiation of the parent strain with UV, causing the creation of mutants different in their spectrum of antimicrobial activity. Treatment of S. fradiae NRRL 2702 with n-nitroso-guanidine (NTG), or exposure to UV formed from tylosin hyper-producing mutants. Mutagenesis by NTG and UV increase the productivity of S. hygroscopicus FC 904 (rapamycin producer) from 60 - 124 % [5].
The objective of the present study is to improve CA production by the Streptomyces sp. MU-NRC77 mutant strain followed by optimization of the fermentation medium and culture conditions.
Methods
Microorganisms and culture conditions
The strain Streptomyces NRC-77 was previously isolated from an Egyptian soil sample by a screening program [6]. This Streptomyces strain produces CA and was deposited in the Actinomycetes Culture Collection, National Research Centre, Dokki, Giza, Egypt. It was maintained in an International Streptomyces Project medium 4 (ISP4) composed of (g/L): Soluble starch, 10.0; K2HPO4, 1.0; MgSO4.7H2O, 1.0; NaCl, 1.0; (NH4)2SO4, 2.0; CaCO3, 2.0; agar, 20.0. A strain of Escherichia coli, maintained in a nutrient agar medium capable of resisting 25 μg/mL of penicillin-G (Glaxo Wellcome UK Ltd., Middlesex, UK), was used as a test strain for evaluation of CA concentration [6]. Colonies of these strains were stored at -80 °C in 20 % glycerol for further studies.
Strain improvement through UV mutagenesis
A sample (10 mL) of spore suspension from a solid culture of Streptomyces sp. NRC-77 was transferred to a sterilized Petri dish plate under aseptic conditions. The plate, with cover removed, was exposed to UV irradiation for different exposure times (30, 60, 90, 120, 150, 180, 210 and 240 seconds) at a distance of 20 cm from the UV lamp with wavelength of 254 nm and power of 30 W. All these exposures were performed in a dark room to avoid any photoreaction in the production of mutants. Spore suspension, exposed or not exposed to UV irradiation, was also spread over the surface of an ISP2 medium for 10 days at 28 oC. Colonies which survived at each dose of the UV irradiation were transferred to ISP2 slants composed of the following (g/L): D (+) glucose, 4; malt extract, 10; yeast extract, 4 and agar, 20, for 10 days. The direct demonstration of CA production was achieved by sub-culturing streaks of each mutant, on plates containing a starch nitrate medium. The plates were then cut off using sterile cork borer and the discs were transferred to Petri dishes containing Muller Hinton agar as a bioassay medium, seeded with a penicillin-resistant test strain of E. coli suspension (OD = 1.0 at 600 nm), then transferred to a refrigerator for 30 min at 4 oC and incubated at 37 oC for 24 h. The diameter of the resulting inhibition zones were then measured and considered to estimate the mutant's CA potentialities [7]. Colonies with a diameter of inhibition zone greater than that of the parent wild type strain Streptomyces sp. NRC-77 were picked up for further secondary screening in shake flasks.
Production medium and cultivation conditions
The most potent selected strain obtained after UV mutagenesis was cultivated in Erlenmeyer flasks (250 mL) containing 50 ml production medium in an Innova 4080 rotary shaker (New Brunswick, NJ, USA) at 200 rpm and 28 °C for six days. The inoculation was conducted using a spore suspension (1.6 × 108 spores/mL) of the most potent strain slanted in an ISP2 medium. The production medium was composed of (in g/l): Soluble starch 25; Glycerol 10 mL; Soybean meal 30; KH2PO4, 0.1, FeSO4 .7H2O, 0.1; and the pH was adjusted to 7.0 before sterilization. Culture broth was separated from the mycelium by centrifugation at 5000 rpm by refrigerated centrifuge (Sigma 3-16 KL laboratory centrifuge, Germany) for 10 min. The supernatant was filtered and used for the evaluation of the inhibitory activity, which was conducted biologically using the agar diffusion method and spectrophotometrically through the reaction of CA with imidazole at 312 nm [7,8].
Fermentation assays were carried out in duplicates. The most potent strain, which showed the greatest CA production, was selected for further investigation.
Determination of CA by high performance liquid chromatography (HPLC)
Clavulanic acid was determined after reacting with an imidazole reagent [8] using SYIKIM HPLC (Germany), a reversed phase column (Polaris C-18, 5 μm, 250 × 4.6 nm), at 30 ºC. The peaks were detected using a variable wave length photos array detector model (Jasko - UV - 2070 Plus, Intelligent UV/Visible detector, Japan). For chromatographic separation, a C-18 column (Thermo Hypersil Keystone, 5 μm, 250 × 4.6 mm) was used. Potassium dihydrogen orthophosphate buffer (pH 3.2) and 6 % methanol was used as a mobile phase and the flow rate was adjusted to 1 mL/min. The sample volume (20 µL) was injected with the help of a micro syringe, the run time was adjusted to 10 min, and UV absorbance was determined at 311 nm. Autochrom 3000 software was the data acquisition system. An authentic sample of CA (Glaxo Wellcome UK Ltd., Middlesex, UK) was used. The results obtained from HPLC analysis of the samples were monitored using the abovementioned authentic sample. The retention time of CA was recorded at 3.4 min [9].
Phenotypic identification
The morphological, cultural, physiological and biochemical characterization of the potential isolate was carried out as described in the International Streptomyces Project (ISP) [10]. The morphology of the spore-bearing hyphae with the entire spore chain, the substrate and aerial mycelium of the strain, was examined by light microscope (1000x magnification) as well as by Transmittance Electron Microscope (TEM). Media used were those recommended in the ISP [11]. Arial mycelium was observed after incubation at 28 °C for 2 weeks and colors series were also observed. Different carbon sources utilization with 1 % were determined by growth in a medium (ISP9) at 28 °C [12]. Nitrate reduction was detected and melanin pigment production was noticed in a medium of ISP6 and 7 [13].
DNA isolation and PCR amplification
An overnight culture of the promised bacteria, grown at 28 ºC, was used for the preparation of genomic DNA. DNA extraction was done by using the protocol of Gene JET genomic DNA purification Kit (Thermo K0721) as following the manufacturer of the kit. The PCR amplification of the 16S rDNA region was carried out following the manufacturer of the Maxima Hot Start PCR Master Mix (Thermo K1051). The 16S rDNA was amplified by a polymerase chain reaction (PCR) using primers designed to amplify 1500 bp fragment of the 16S rDNA region. The domain bacteria-specific primer 27F (forward primer) was 5’AGAGTTTGATCMTGGCTCAG3’ and the universal bacterial primer 1492R (reverse primer) was 5’TACGGYTACCTTGTTACGACTT3’ [14].
The PCR reaction was performed with 5μl of genomic DNA as the template, 1μl of 16S rRNA Forward primer, 1 μL of 16S rRNA reverse primer, 18 μL nuclease-free water, and 25 μL Maxima® Hot Start PCR Master Mix (2X) in a 50μL reaction mixture as follows: activation of 2 Taq polymerase at 95 °C for 2 min; 35 cycles of 95 °C for 1 min, 65 °C, and 72 °C for 1 min each were performed, finishing with a 10-minute step at 72 °C. After completion, the PCR products were electrophoresed on 1 % agarose gels, containing ethidium bromide (10 mg/mL) [15], to ensure that a fragment of the correct size had been amplified.
DNA sequencing, phylogenic analysis and tree construction
The amplification products were purified with a K0701 Gene JET™ PCR Purification Kit (Thermo). Afterward, the samples become ready for sequencing in an ABI Prism 3730XL DNA sequencer and analyzer from the GATC Company. The sequencing reaction was performed with the primers 518F 5' (CCA GCA GCC GCG GTA ATA CG) 3' and 800R 5' (TAC CAG GGT ATC TAA TCC) 3', using a PRISM BigDye Terminator v3.1 Cycle sequencing Kit. The DNA samples containing the extension products were added to Hi-Diformamide (Applied Biosystems, Foster City, CA). The mixture was incubated at 95 °C for 5 min, followed by 5 min on ice and then analyzed by an ABI Prism 3730XL DNA analyzer (Applied Biosystems, Foster City, CA). The sequence alignment was prepared with DNASTAR software programs (DNASTAR. INC., Madison,Wis.).
Phylogenetic data were obtained by aligning the nucleotides of different 16S RNA retrieved from a BLAST algorithm (www.ncbi.nlm.nih.gov/ BLAST), using the CLUSTAL W program version 1.8 with standard parameters. The classifier is trained in the new phylogenetically consistent higher-order bacterial taxonomy (Ribosomal Database Project, RDP Classifier) previously proposed [16] (http://rdp.cme.msu.edu/classifier/ classifier.jsp).
Phylogenetic and molecular evolutionary analyses were conducted using Mega6. A rooted phylogram was derived from the distance matrices using the neighbour-joining method through the Mega 6. All analyses were performed on a bootstrapped data set containing 1000 replicates (generated by the program).
Nucleotide sequence ID
The nucleotide sequence of the 16S rRNA gene of strain NRC-77 has been deposited in Gen Bank under the ID: KT953342.
Fermentation medium optimization
The effect of different incubation periods on cell growth and CA production was studied. To determine the best carbon source that provides the maximum yield of CA by the most potent mutant strain, each of the carbon sources of native starch, raw rice powder, and glycerol, were added instead of soluble starch in the fermentation medium.
The effect of the combination of the most favorable carbon source was investigated. On the other hand, the effect of different inoculum types, sizes and some additives were also investigated.
Determination of the cell dry weight
Fermentation culture samples were filtered through a pre-weighed filter paper (Whatman No. 1) and washed four times with 20 mL distilled water. Wet filter papers were dried overnight at 70 oC and then weighed after cooling in a desiccator. The dry weight in grams per liter from the fermentative culture was determined.
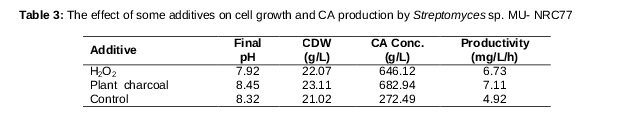
Effect of hydrogen peroxide and activated animal charcoal on cell growth and CA production by Streptomyces sp. MU-NRC77
Redox cyclic agents such as hydrogen peroxide (H2O2) were added at a concentration of 10 µM, in dimethylsulfoxide (DMSO) at a final concentration of 1 % after 38 h of incubation. It was previously sterilized by a 0.22 µm Millipore syringe filter [17]. In the case of heat-sterilized activated animal charcoal, it was added to the production medium at 1.0 % before inoculation, according to Ivanitskaya et al [18]. All chemicals used were in the analytical grade.
Results
Strain inprovement by UV radiation
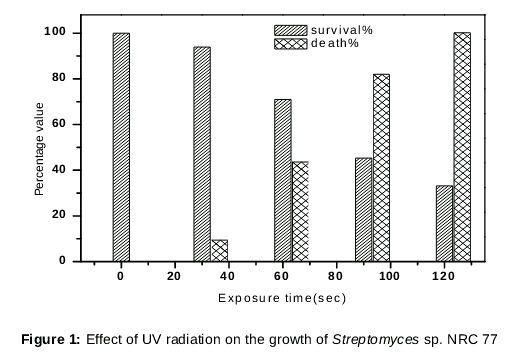
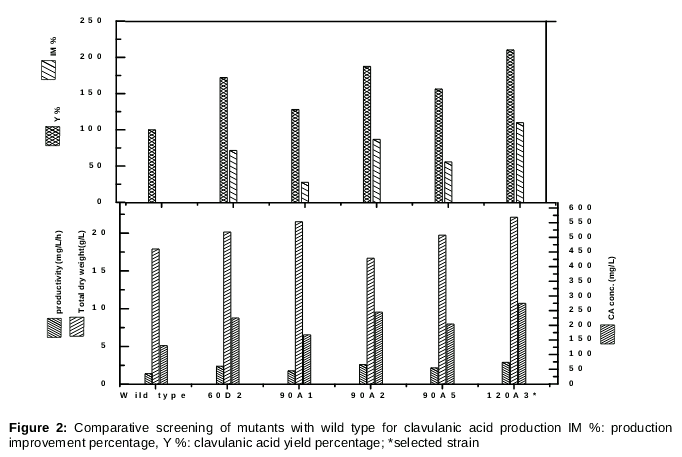
To improve the production of CA, the Streptomyces sp. NRC-77 wild-type strain was exposed to the distinctly potent mutagen UV light. After 10 days of incubation, colonies of mutants grown on ISP2 were obviously observed. The considerable variability observed between the surviving isolates from each UV dose, pointed out the strong mutagenic effect of the UV radiation. Mutant colonies exposed to UV radiation for 30 seconds showed a decrease in total count with increasing time of exposure. The lethality percentage increased from 6 to 66.8 % with exposure time elevation from 30 to 120 seconds radiation as shown in . After the exposure of UV irradiation, CA was improved and higher yield was detected from the mutant strains compared with the parent strain. Mutant 120A3 showed the highest CA yield reaching 275 mg/L compared with the wild strain (NRC77), which reached 130.70 mg/L [6], and with the other mutant strains as shown in .
An increase in productivity of 47.5 more than the parent strain was recorded by the most potent mutant. In our current study, the genetic constancy of the most potent mutant Streptomyces sp. MU-NRC77 strain was examined by multiple sub-culturing in a starch nitrate medium and then analyzing the production of the desired CA by agar diffusion, spectrophotometry and confirming by the HPLC method. Consequently, the mutant 120A3 was subjected to identification.
Characteristics and identity of potential isolate strain, 120a3
Phenotypic characteristics
Under microscopic observation, strain 120A3 showed the presence of both long aerial mycelium, not fragmented, with open spiral spore chains and substrate mycelium with smooth spore chains as shown in .
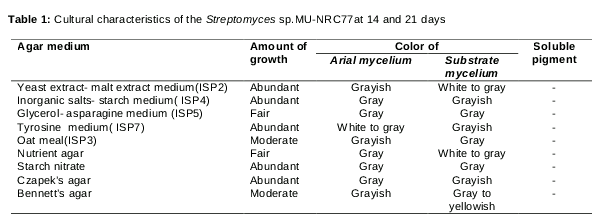
The morphology of the isolate resembled Streptomyces with its slow growing colonies, aerobic, glabrous or chalky and folded aerial mycelium. In addition, it possessed an earthy odor. The mutant isolate showed good growth in (ISP2, ISP4 and ISP7) media. Moderate and poor growth was observed in ISP3 and ISP5 media respectively. The color of aerial mycelium was subjected to the gray series ().
The isolate reduced nitrate utilized a wide range of sugars as a carbon source and did not produce diffusible melanin pigments as shown in .
Based on its phenotypic properties, strain MU-NRC77 was classified in the genus Streptomyces. The characteristics of this strain were compared with the known Streptomyces species in Bergey’s ‘Manual of Systematic Bacteriology [13]. Strain MU-NRC77 has a similarity to S. rochei and the strain NRC-77 wild type. These strains have the same aerial mycelium colors, spore shapes and physiological characteristics, although with certain differences between them. Strain MU-NRC77 differs from all of the previously listed Streptomyces species in its characteristics, such as its utilization of D (+) glucose, cellobiose and cellulose and the fact that it produces CA, unlike other species.
Genotypic characteristics
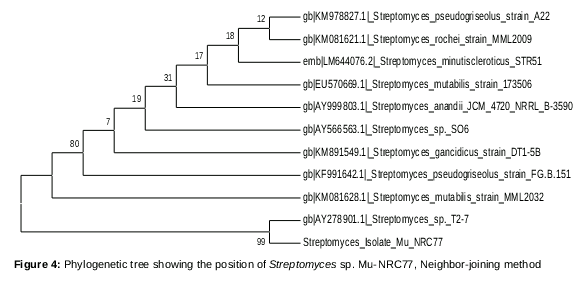
The phylogenetic tree () of Streptomyces sp. MU-NRC77 was constructed based on its 16S rRNA gene sequence, which has been submitted to GenBank under accession number (ID: KT953342).
In the phylogenetic tree (), strain MU-NRC77 was posed with Streptomyces sp.T2-7(gb/AY278901.1) in a branch. Streptomyces mutabilis strain MML2032 (gb/KM081628.1) was found to be the neighbor strain. Streptomyces sp. MU-NRC77 was posed with Streptomyces sp.T2-7(gb/AY278901.1) as a single branch, sharing 99% sequence similarity and supported by bootstrap value 99. It is clear from the phylogenetic analysis that strain MU-NRC77 is suggested to be a new variety of Streptomyces rochei. Thus, the newly isolated strain MU-NRC77 is designated Streptomyces sp. MU-NRC77.
Optimization of fermentation medium
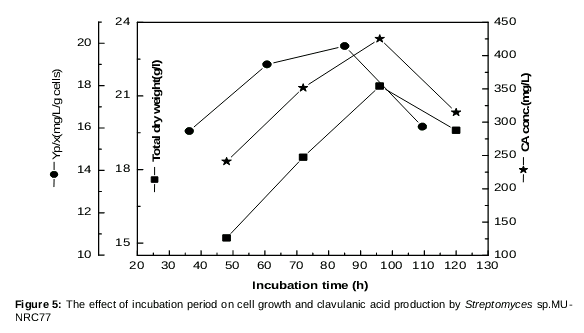
Effect of different incubation period on cell growth and CA production
The analysis of results in showed that CA was increased linearly up to 96 h, giving the maximum yield of 275 mg/L and yield coefficient (Yp/x) of 12.85 mg/g cells. Above this phase of growth, there was a slight decrease in antibiotic production as the incubation time reached 250 mg/L at 120 h. The final pH moved towards alkalinity during the fermentation period, reaching a pH value of 8.4 at the optimum time; a further increase in pH was noticed, reaching its highest value of 8.6 at 120 h (data not shown).
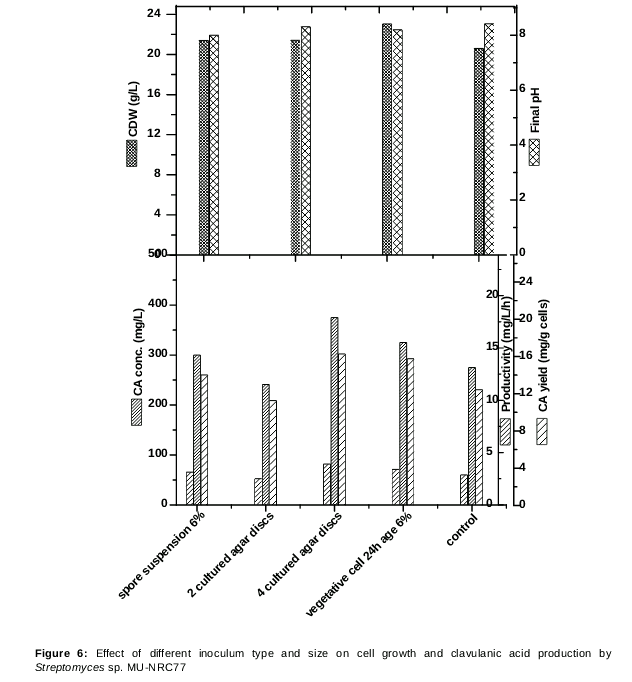
Effect of different inoculum type and size on cell growth and CA production by Streptomyces sp. Mu-nrc77
The effect of inoculum type and size was also performed. In this case, inoculation with spore suspension was favored in 6 % of vegetative cell at 24 h age per 50 mL production medium of both. Also, it was noticed that the CA yield obtained by inoculation of spores discs cultured in an agar medium supported higher production than spore suspension type. Furthermore, maximum production of 375 mg/L was achieved using an inoculum size of 4 discs of cultured agar medium. Also, results showed that generally, the cell growth dry weight increased by increasing the inoculum size. These results revealed a parallel increase in CA production by using 4 discs of cultured agar medium rather than 2 discs as shown in .
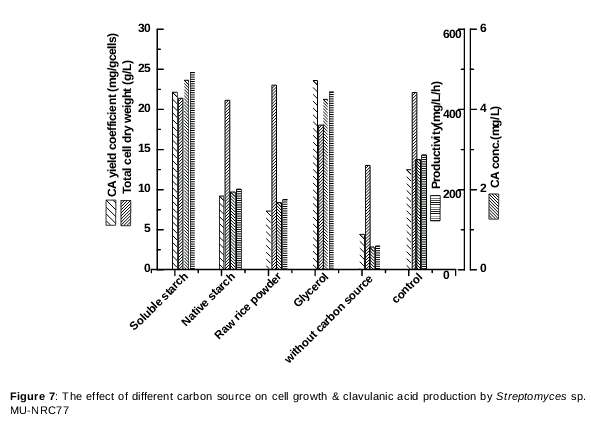
Effect of different carbon sources and combination with soluble starch: glycerol
The ability of the mutant strain Streptomyces sp.MU-NRC77 to grow and produce CA with different carbon sources containing media was tested and compared with a medium without a carbon source. Out of the five tested carbon sources, only soluble starch and glycerol supported good CA production (475 and 450 mg/L). As well as this, the production yield YP/x recorded the highest values, underlining the advantage of using glycerol and soluble starch as carbon sources in the fermentation medium, whereas a medium without a carbon source did not support CA production at any measurable level, as shown in .
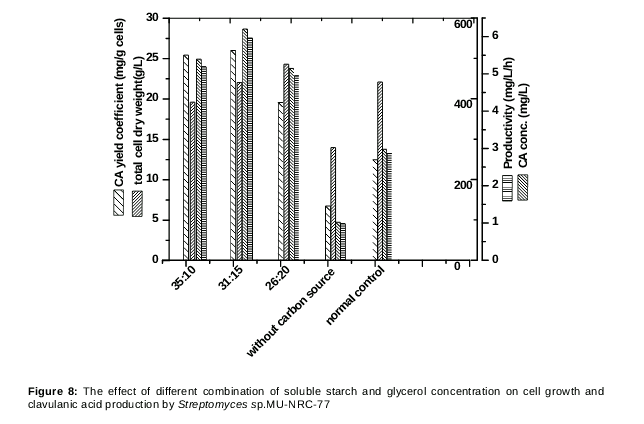
Various ratios of soluble starch in combination with glycerol were examined for improving the CA production. As shown in , soluble starch in combination with glycerol by 31:15 w/v gave higher CA production (572 mg/L) than that obtained when each one was used alone in the fermentation medium. The biomass concentration showed a slight increase with the medium containing higher glycerol concentration in combination with starch. Consequently, these proportions were used in the final optimized fermentation medium as follows (g/L): soluble starch 32.0; glycerol, 15 mL; soybean meal, 30.0; KH2PO4, 0.1; FeSO4.7H2O, 0.1; agar 15; Trace elements solution, 1 (mL/L) FeSO4.7H2O, 1; MnCl2.4H2O, 1; ZnSO4.7H2O, 1 and CaCl2, 1 and pH 6.8 [19].
Effect of hydrogen peroxide and activated animal charcoal on cell growth and CA production by Streptomyces sp. MU-NRC77
The results in indicated that culture media supplemented with H2O2 enhanced CA production, with a recorded CA yield of 646.12 mg/L with a productivity of 6.73 mg/L/h, compared to that obtained from the control culture 275 mg/L with a productivity of 4.92 mg/L/h. On the other hand, the maximal CA yield 683 mg/L with a productivity of 7.11 mg/L/h was listed using activated animal charcoal, which revealed an improvement in CA yield by 42.56 and 40.26 % respectively.
Discussion
The production of secondary metabolites by improved strains through mutagenesis techniques is considered an advantage during the fermentation process [20]. Random mutation by UV irradiation is a design for improving strains towards higher production [21]. The antitumor glycopeptide antibiotic zorbamycin produced from Streptomyces flavoviridis ATCC 21892 was enhanced through strain improvement by using UV radiation [22]. In another study, mutation and screening of the parent culture to improve rapamycin production were carried out [23]. It was reported previously that the lethality rate should be extremely high in order to have powerful mutations and effective screening of mutants.
As a result of UV mutagenesis when UV exposure time was increased from 0 to 25 minutes, a decrease in the survival rate of S. venezuelae from 100 % to 8 % occurred. It is noteworthy that a gradual reduction in survival rates was observed by the increase in UV exposure time on S. fradiae NRRL-2702. An enhancement in tylosin production by the resultant mutants compared to the wild type strains was detected [20].
Several authors have reported that the 16S rRNA gene sequence has been demonstrated to be a powerful method for investigating the phylogeny of microorganisms and a tool for analysis and differentiation (differentiation unknown at the species or strain level) of the genus Streptomyces and can be used as a genetic method in parallel to actinomycetes’ conventional taxonomic methods [24]. In this work, we demonstrate that the mutant isolate formed a distinct phyletic line in the16S rRNA gene tree and has chemotaxonomic properties typical of the genus Streptomyces and was closely related to Streptomyces sp.T2-7. It was accordingly given the name of type strain Streptomyces sp. MU-NRC77.
A series of experiments was conducted to determine the optimal physiological and fermentation conditions leading to the improvement of clavulanic acid production by the experimental microorganism. The time course showed that CA began to be produced from the first day and reached a maximum on day four. The instant decrease in CA yields upon attaining their highest value has not been explained yet, although it is an obvious behavior in all trials [21].
The production rate remained almost constant from 72 h up to 96 h, possibly after carbon sources consumption had been completed. Also cell growth indicated that production of CA is associated with Streptomyces sp. MU-NRC77 cell growth, the results showing that CA rate production diminished after 96 hours of cultivation. This may be due to the inhibition and repression activity of a product from the metabolism of the microorganism acting on the metabolic pathway of CA production [1]. Production of antimicrobial metabolites from microorganisms requires glycerol as a known important medium component [25]. Also, glycerol supplies the carbon skeleton of the ß-lactam ring without conformational change in the three carbon positions [26] In our findings, CA production was higher using glycerol as sole carbon source, in agreement with earlier work reported by Bellao et al [27].
It has previously been reported that slowly used complex carbohydrates catalyze the production of secondary metabolites produced by actinomycetes in the production media, therefore, native starch, and raw rice powder with soluble starch were tested individually as possible carbon sources. Therefore, substitution of soluble starch and glycerol in the production media with native starch and raw rice powder as sources of carbon resulted in diminishing CA production. The cost benefit ratio of these carbon sources is important for the growth and metabolism of the producer microorganism. Therefore, glycerol and soluble starch were chosen as the best carbon sources and their combination ratio of 31:15 improved production. It is well known that inoculum type and size strongly influence the efficiency of microbial culture. The inoculum size was important for metabolite production as previously mentioned. Lower inoculum size needs prolonged time for cells to multiplication to a high enough number for utilizing the substrate and producing CA.
The biomass regeneration and synthesis were realized by the increase in inoculum cell number. This fact was supported by our findings as the strain showed a parallel increase in clavulanic acid with the increase in inoculum size [25]. Previous reports showed that the seed stage manipulation improved rapamycin production by Streptomyces hygroscopicus. When inoculum size was 6 % the highest rapamycin yield was attained [28].
It is believed that metabolic stimulus may increase CA production. A culture that contained H2O2 increased the CA yield by 2.34-fold, which is in agreement with results obtained by Kwon and Kim [17].The maximal CA yield obtained from adding activated animal charcoal to the optimized medium increased the CA yield by 2.48 fold. This finding was in accordance with a previously mentioned report which studied the effect of activated charcoal on the CA synthesis by Streptomyces clavuligerus, proving that the CA synthesis rate and yield were increased 27. Moreover, they detected CA within 24 h in a medium containing 1 % charcoal and the CA yield increased at least 4-fold and reached its maximal after 72 h instead of 96 hours.
Conclusion
A mutant strain, Streptomyces sp. MU-NRC77, capable of showing hyper-production of CA, has been successfully isolated via UV irradiation mutation, followed by medium optimization. An increase in CA yield of approximately 5.2 fold compared to the wild type strain was obtained. Planning toward the application of large scale production of CA in a bioreactor by the mutant strain will explore the economic outcome of mutagenesis and optimization studies in our work.
Declarations
Acknowledgement
References
Archives


News Updates